はじめに
今回は自然免疫と獲得免疫について詳しく解説していきます!免疫反応の本質を理解していただくために、詳細な情報はできるだけ省略していますが、重要な部分はしっかりと解説しています。また、「免疫学に関するFAQ」項では、私が免疫学を勉強していた時に実際に疑問に思ったことをQ&A方式で解説しています。皆様もおそらく同じ箇所について疑問に感じると思いますので、きっと役に立つと思います。ぜひ最後までご覧ください。
免疫学は複雑で難しいです。複雑だからこそ美しく、非常に面白い学問分野であると感じています。読者の皆様にも、その美しさや面白さを感じていただけたら幸いです。
難しい部分もあるかとは思いますが、じっくりと読み進めてください!
【難易度】★★★★★
【重要度】★★★★★
※本記事で登場する免疫細胞がどのように体を循環しているか?免疫反応はどこで起きているか?について以下の記事で詳しく解説しています。ぜひご覧ください!
免疫系の概要
自然免疫と適応免疫の細かい内容に入る前にまずは、ウイルスが体内に侵入してきた時を例に、免疫系がどのように機能するかを順番に解説していきます。 細かい内容は次項で解説していますので、ここでは大体の流れを把握できれば大丈夫です。また、次項以降を読み進めているうちに頭が混乱してきたら、本項に戻って全体の流れの中のどこの部分の話をしているかを確認しましょう。
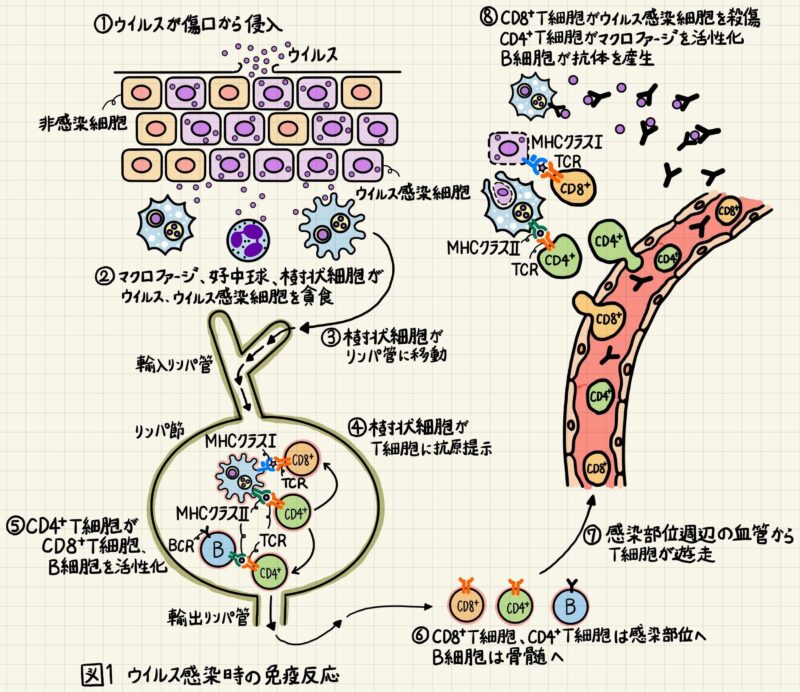
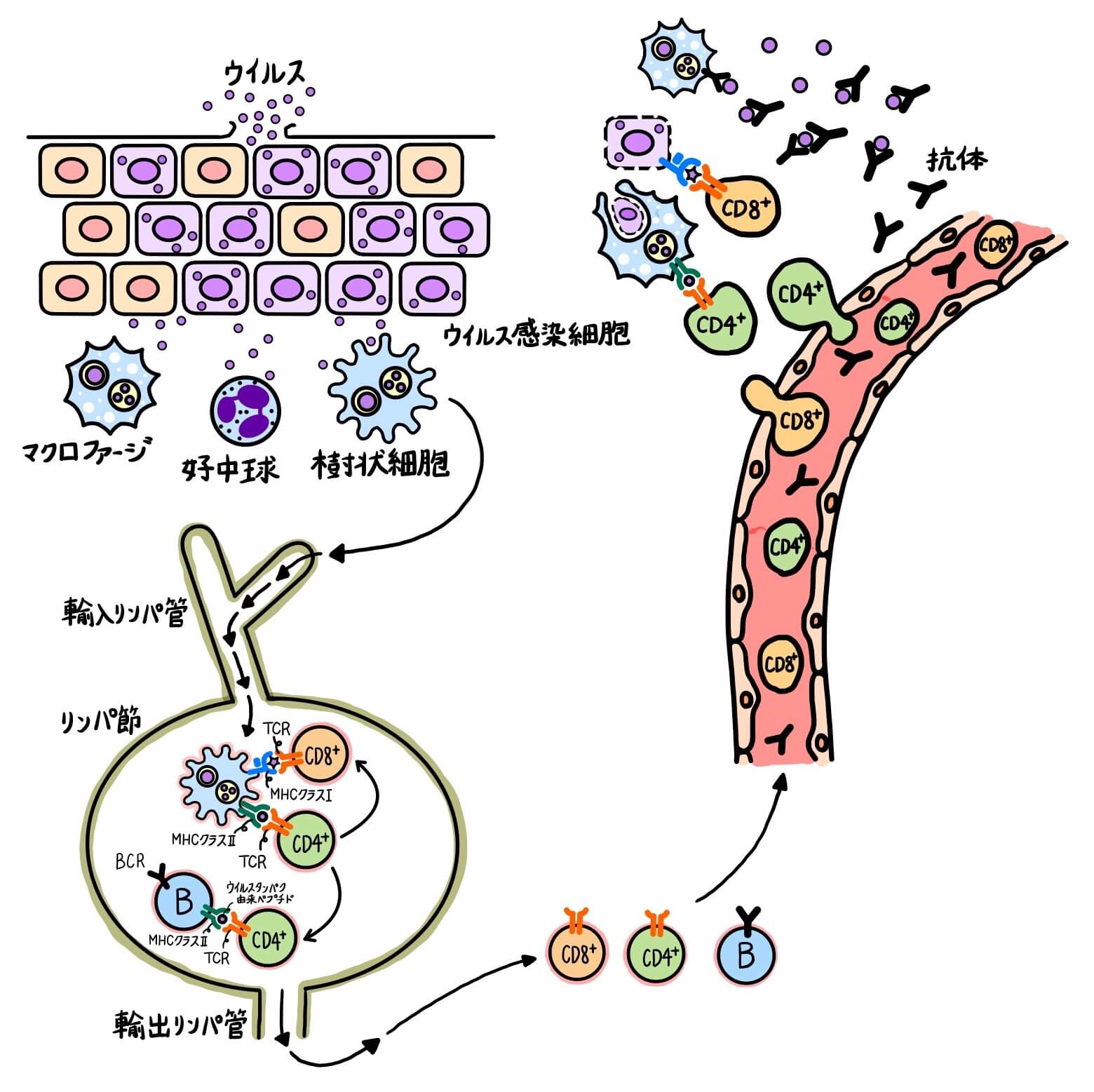
【図1①, ②】ケガをして病原体(ウイルス、細菌、菌類など)が体内に侵入した時、まず好中球、マクロファージ、樹状細胞などの食細胞がウイルスを素早く食胞(ファゴソーム)に取り込みます。その後、ファゴソームは酸性加水分解酵素などが多く含まれるリソソームと融合し、ウイルスはペプチド断片まで分解されます。こうしてウイルスは体内から排除されます。この好中球とマクロファージを中心とした強力な生体防御システムを自然免疫と言います。自然免疫系の細胞は、病原体に共通して存在する分子パターンを認識して、素早く病原体をファゴソーム内に取り込みます。
分解されたウイルスタンパク由来のペプチド断片は細胞内でMHCと呼ばれるタンパク質と結合し、細胞表面に提示されます。 MHC(major histocompatibility complex; 主要組織適合複合体)は、細胞膜表面にある糖タンパク質のことで、クラスIとクラスⅡの2種類があります。MHCクラスⅠは赤血球以外の全ての細胞で発現していますが、MHCクラスⅡは抗原提示細胞(樹状細胞、マクロファージ、B細胞)と呼ばれる細胞種でのみ発現しています。
【図1③, ④, ⑤】ウイルスを取り込んで活性化した樹状細胞はリンパ節に移動し、「MHCクラスⅡ+ウイルスタンパク由来ペプチド」をナイーブCD8+T細胞とナイーブCD4+T細胞に提示します(抗原提示)。樹状細胞が提示した「MHCクラスⅡ+ウイルスタンパク由来ペプチド」を認識できるナイーブCD8+T細胞とナイーブCD4+T細胞は増殖・分化してエフェクター細胞になります。
【図1⑥, ⑦】エフェクターCD8+T細胞の大部分はウイルス感染部位周辺の血管から感染部位に移動して、ウイルスに感染した細胞を殺します。ウイルスに感染した細胞は「MHCクラスI+ウイルスタンパク由来ペプチド」を細胞表面に発現しており、CD8+T細胞はこの「MHCクラスI+ウイルスタンパク由来ペプチド」を識別します。エフェクターCD4+T細胞は、B細胞やマクロファージが発現している「MHCクラスⅡ+ウイルスタンパク由来ペプチド」を識別してB細胞、マクロファージを活性化します。エフェクターB細胞は形質細胞に分化し、抗体を産生します。抗体はウイルスが細胞に結合するのを防いだり、食細胞や補体系を活性化したりします。マクロファージとB細胞も「MHCクラスⅡ+ウイルスタンパク由来ペプチド」を発現する抗原提示細胞ですが、ナイーブT細胞・ナイーブB細胞を活性化し、適応免疫を誘導するのは原則樹状細胞です。
T細胞とB細胞を中心とする生体防御システムを適応免疫と言います。適応免疫系は発動に数日要しますが、病原体特異的で非常に強力です。適応免疫系が発動するまでの数日間は自然免疫系のみで病原体と戦っています。
ヒトのMHCをHLA(Human leukocyte antigen; ヒト白血球抗原)と言います。ほとんど全ての細胞に発現しているのにも関わらず、「白血球」という名前が入っているのは白血球に特異的な細胞表面分子として発見されたからです。ちなみに、マウスにおけるMHCはH-2(histocompatibility-2)と言います。
図2は図1を簡略化して描いた図です。分かりやすい方を復習に使って頂けたら幸いです。
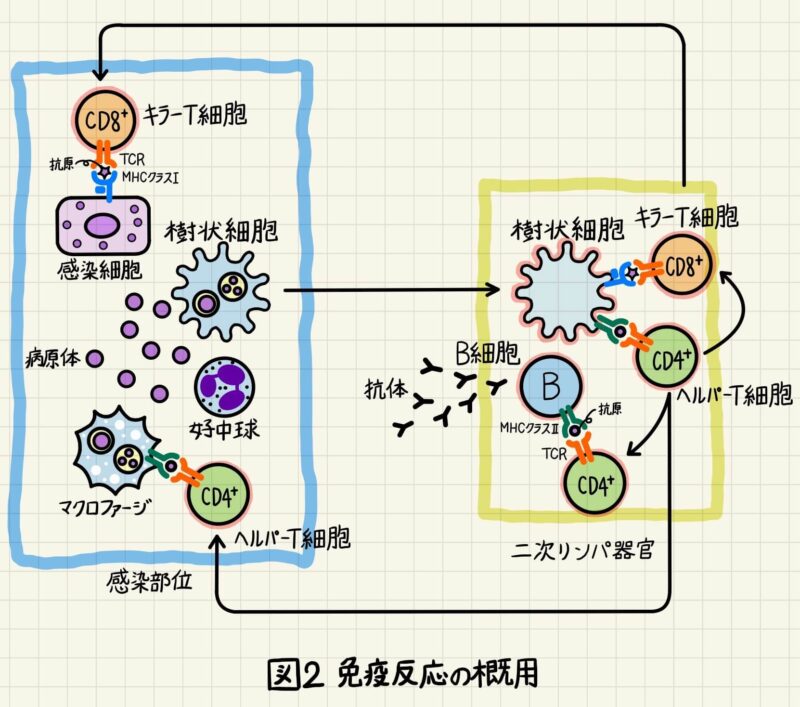
自然免疫系と適応免疫系の相互作用
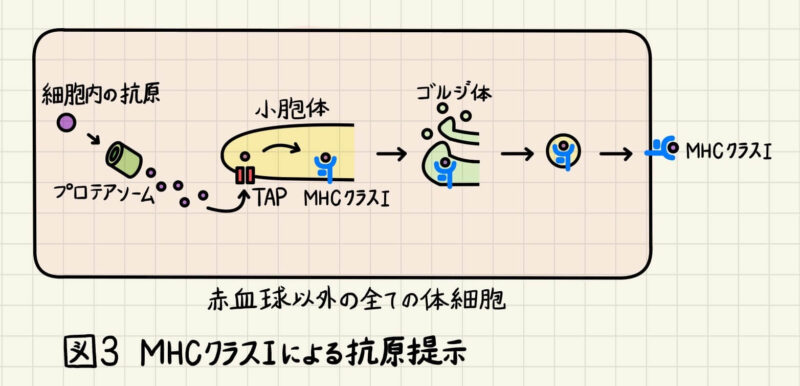
MHCクラスIによる抗原提示
細胞質に存在する一部のタンパク質はプロテアソームによって分解され、TAP(transporter associated with antigen presentation)と呼ばれるトランスポーターを通って小胞体に入ります。小胞体に入ったペプチド断片はMHCクラスIと結合し、細胞表面に運ばれます(図3)。例えば、細胞にウイルスが感染した時、上述の仕組みで「MHCクラスⅠ+ウイルスタンパク由来ペプチド」が細胞表面に提示されます。これがウイルスに感染した細胞であることの目印になり、CD8+T細胞(=キラーT細胞)が感染細胞を殺傷します。 T細胞は、「MHC+ペプチド」をT細胞受容体(TCR; T-cell receptor)を使って認識します。ここで重要なことは、TCRはペプチドだけに結合するのではなく、MHCタンパクとも結合するということです。
また、もう一つ重要なことは、MHCクラスⅠによって細胞表面に提示されるのは、病原体由来のペプチド断片だけでないということです。「MHCクラスⅠ+自己タンパク由来のペプチド断片」も大量に細胞表面に提示されています。自己タンパク由来のペプチドを認識するT細胞は、T細胞分化の過程で排除されるので、自己の細胞を攻撃してしまうことはありません。
自己タンパク由来のペプチドを非自己だと勘違いしてT細胞が攻撃してしまう病気があります。それが各種アレルギー疾患や、間接リウマチ、I型糖尿病、多発性硬化症などの自己免疫疾患です。
CD8はキラーT細胞特異的に発現しているタンパク質のこと。CD8+T細胞は「MHCクラスⅠ+ペプチド」を認識して、病原体に感染した細胞を殺傷する。
MHCクラスⅡによる抗原提示
マクロファージや樹状細胞などの食細胞は、細胞外の病原体を細胞内に取り込んで分解します。分解されたペプチドはMHCクラスⅡと結合し、細胞表面に運ばれます(図4)。細胞外タンパク由来のペプチドとMHCクラスⅡが結合するまでの間、MHCクラスⅡと細胞質内タンパク由来ペプチドが結合しないように、インバリアント鎖と呼ばれるCLIPがMHCクラスⅡに結合しています(図4)。例えばウイルスを飲み込み、「MHCクラスⅡ+ウイルスタンパク由来ペプチド」を発現した樹状細胞はリンパ節に移動し、ナイーブCD4+T細胞(ヘルパーT細胞)に抗原提示します(図1)。抗原提示により活性化したCD4+T細胞は、「MHCクラスⅡ+ペプチド」を提示したB細胞、マクロファージを活性化します。詳細は解明されていませんが、CD4+T細胞は CD8+T細胞のT細胞の活性化にも関与しています。
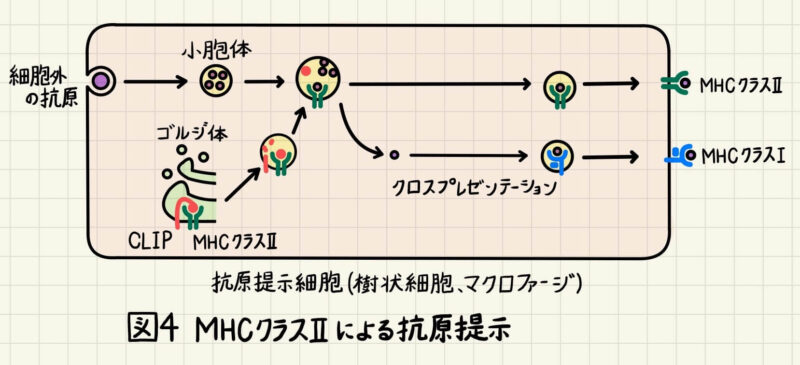
また、細胞外から取り込まれた抗原は必ずしもMHCクラスⅡに積み込まれるわけではなく、細胞質に放出されてプロテアソームで分解されるものがあります。そして、前項に記述した方法で「MHCクラスⅠ+ペプチド」としても抗原提示細胞の細胞表面に提示されます。これをクロスプレゼンテーションと言います(図4)。クロスプレゼンテーションにより、抗原提示細胞は「MHCクラスⅠ+細胞外タンパク由来ペプチド」を細胞表面に提示することができます。
例えばウイルスを飲み込み、「MHCクラスⅠ+ウイルスタンパク由来ペプチド」を発現した樹状細胞はリンパ節で、CD8+T細胞(キラーT細胞)に抗原提示します。抗原提示により活性化したCD8+T細胞は、「MHCクラスⅠ+ウイルスタンパク由来ペプチド」を提示したウイルス感染細胞を殺傷します。
CD4はヘルパーT細胞特異的に発現しているタンパク質のこと。CD4+T細胞はCD8+T細胞、B細胞、食細胞の活性化を行う。
ウイルスに感染した全ての細胞は、「MHCクラスⅠ+ウイルスタンパク由来ペプチド」を細胞表面に提示して、感染したことをCD8+T細胞に知らせます。
一方で、抗原提示細胞は「MHCクラスⅡ+ウイルスタンパク由来ペプチド」と「MHCクラスⅠ+ウイルスタンパク由来ペプチド」を細胞表面に提示します。「MHCクラスⅡ+ウイルスタンパク由来ペプチド」はCD4+T細胞を、「MHCクラスⅠ+ウイルスタンパク由来ペプチド」はCD8+T細胞を活性化します。
免疫学に関するFAQ
CD8+T細胞はウイルス感染細胞と抗原提示している樹状細胞をどのように見分けているのか?
CD8+T細胞は「MHCクラスⅠ+ウイルスタンパク由来ペプチド」を発現している樹状細胞により活性化され、「MHCクラスⅠ+ウイルスタンパク由来ペプチド」を発現しているウイルス感染細胞を殺傷します。つまり、CD8+T細胞はTCRを介して結合した相手が樹状細胞か、ウイルス感染細胞か分からないということです。 実際にエフェクターCD8+T細胞は、「MHCクラスⅠ+ウイルスタンパク由来ペプチド」を細胞表面に提示した樹状細胞に出会うと、樹状細胞を殺してしまいます。せっかく適応免疫を発動させようと抗原提示している樹状細胞を、CD8+T細胞が殺してしまったらもったいないですね。しかし、この一見無駄とも思える樹状細胞の排除は、炎症の収束に必要だと考えられます。また、樹状細胞もウイルスに感染することがあるので、もし仮にCD8+T細胞が樹状細胞を殺せないとなると、ウイルスに感染した樹状細胞を体内から排除できなくなってしまいます。
MHCクラスⅠ+ウイルスタンパク由来ペプチド」を発現したウイルス感染細胞はCD8+T細胞を活性化できるか?
樹状細胞はクロスプレゼンテーションにより、細胞外から取り込んだペプチドをMHCクラスⅠとともに細胞表面に提示します(図1)。ウイルス感染細胞も「MHCクラスⅠ+ウイルスタンパク由来ペプチド」を細胞表面に提示しています。図1を見ると、ウイルス感染細胞もCD8+T細胞に抗原提示できそうだと思ってしまいます。しかし、これは起こりません。
なぜならば、T細胞の活性化には抗原提示細胞特有の細胞表面タンパクであるCD80/CD86(共刺激分子)や、抗原提示細胞から産生される各種サイトカインなどが必要だからです。抗原提示細胞以外の細胞にはこれらの機能はないので、CD8+T細胞を活性化することはできません。
また、ナイーブT細胞が末梢組織に移動することも、抹消で感染した細胞が二次リンパ器官に移動することもないので、そもそもウイルス感染細胞とナイーブT細胞は出会いません。適応免疫の始まりの場所である二次リンパ器官に抗原を運んでくれるのは樹状細胞だけなのです。
抗原に遭遇したことのないT細胞のこと。特異的な抗原に出会うと増殖・分化してエフェクターT細胞になる。エフェクターT細胞はウイルス感染細胞を殺したり、他の免疫細胞を活性化したりする。
B細胞はナイーブCD4+T細胞を活性化できるか?
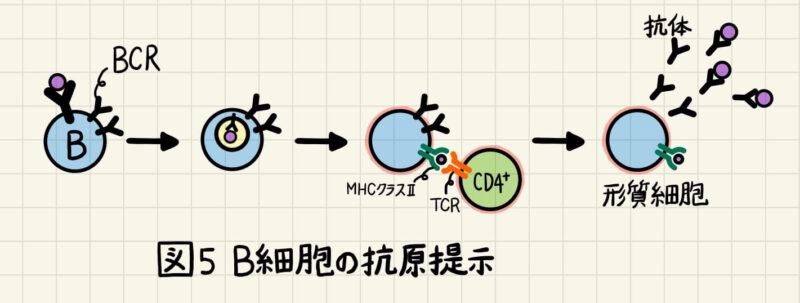
B細胞は、B細胞受容体(BCR; B-cell receptor)に結合した病原体を細胞内に取り込みます。取り込まれた病原体はペプチド断片まで分解され、MHCクラスⅡと結合して細胞表面に運ばれます (図5)。CD4+T細胞はB細胞の細胞表面にある「MHCクラスⅡ+ペプチド」を認識し、B細胞を活性化させます。活性化したB細胞は形質細胞へと分化し、BCRと同じ抗原結合部位を持った抗体を産生するようになります。
樹状細胞やマクロファージなどの食細胞は病原体をどんどん取り込み、多種多様な抗原をT細胞に抗原提示します。一方でB細胞はBCRに結合した病原体を効率的に取り込みます。抗原特異的なBCRを持つB細胞の頻度は非常に少なく、同じ抗原に特異的なナイーブCD4+T細胞と出会う確率がかなり低いので、T細胞の活性化には関与できないと考えられています。
一方で、なぜCD4+T細胞が数少ない抗原特異的B細胞を活性化できるかというと、樹状細胞により活性化したCD4+T細胞は増殖していて、抗原特異的B細胞に出会いやすくなっているからです。
マクロファージはナイーブCD4+T細胞を活性化できるか?
マクロファージは樹状細胞と同様に、ナイーブCD4+T細胞を活性化することができます。しかし、樹状細胞と比較するとその効率は低いです。なぜならば、樹状細胞は好中球やマクロファージなどの食細胞と比較してファゴソームの分解活性がはるかに低く、安定して抗原提示することができるからです。一方で、マクロファージは分解活性が高ために、安定した抗原提示機能が損なわれているようです。また、樹状細胞と異なり、組織中のマクロファージは病原体を取り込んでもリンパ節に移動しないため、ナイーブCD4+T細胞の活性化への寄与は小さいです。
まとめ
- 病原体が体内に侵入した時、食細胞(好中球、マクロファージ、樹状細胞)が病原体を飲み込んで排除する。これを自然免疫という。
- 樹状細胞はCD4+T細胞、CD8+T細胞を活性化して適応免疫を誘導する。
- CD4+T細胞はマクロファージ、B細胞、CD8+T細胞を活性化する。CD8+T細胞は病原体に感染した細胞を殺傷する。
- MHCクラスⅠは赤血球以外の全ての細胞に発現しており、細胞質内のペプチドを細胞表面に提示する。
- MHCクラスⅡは抗原提示細胞(B細胞、マクロファージ、樹状細胞)にのみ発現しており、細胞外のペプチドを細胞表面に提示する。
- 抗原提示細胞は細胞外のペプチドをMHCクラスⅠとともに細胞表面に提示することができる(クロスプレゼンテーション)。
- Savina A, Amigorena S. Phagocytosis and antigen presentation in dendritic cells. Immunol Rev. 2007 Oct;219:143-56. doi: 10.1111/j.1600-065X.2007.00552.x. PMID: 17850487.